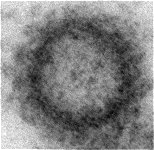
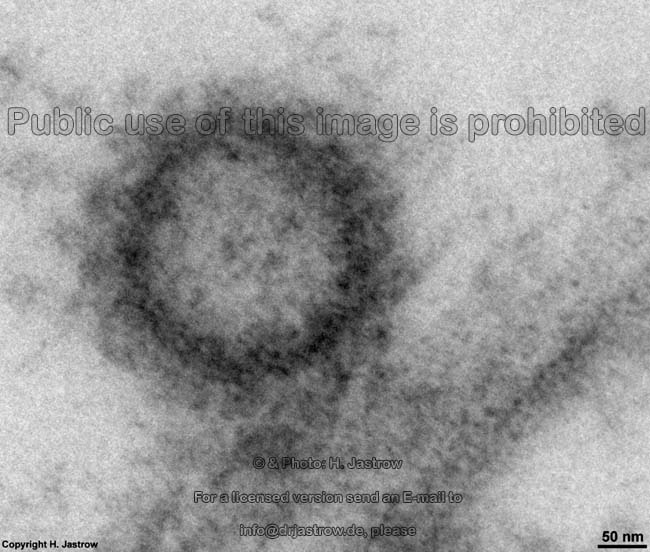
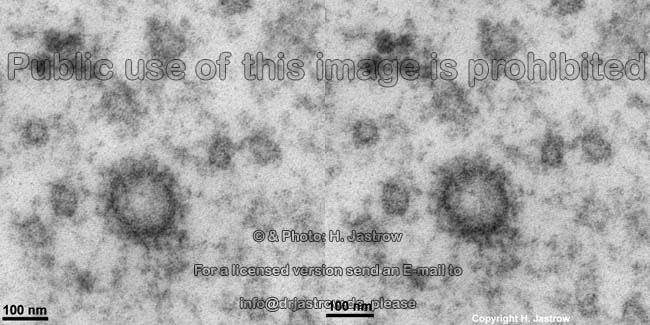
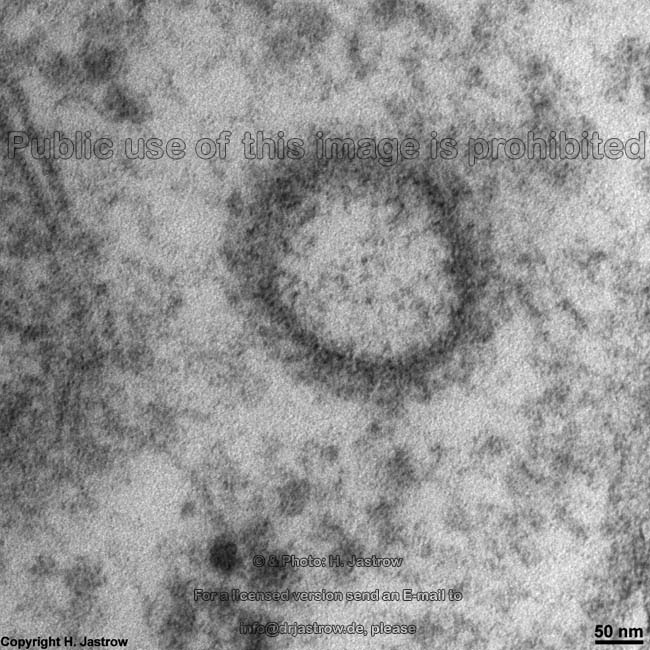
List of abbreviations
of micros-
copic
anatomy
specialist terms
explained in
English +
German

Every attempt was made to provide correct information and labelling, however any liability for eventual errors or incompleteness is rejected!

dieser Seite
Editor:
Dr. med.
H.
Jastrow

Conditions
of use