Fig.1
 |
Ribbon synapses are typical for sensory organs that have a high frequency of output like the retina. These signal transduction foci are characterized by pre-synaptic ribbons (SR; Fig.2). The proteinaceous core of these tiny organelles binds glutamate containing transmitter vesicles. Earlier investigations of the retina showed light dependent morphological changes of SR which were most striking in BALB/c mice. Therefore we chose this strain for study of their three dimensional morphology. | Fig.2
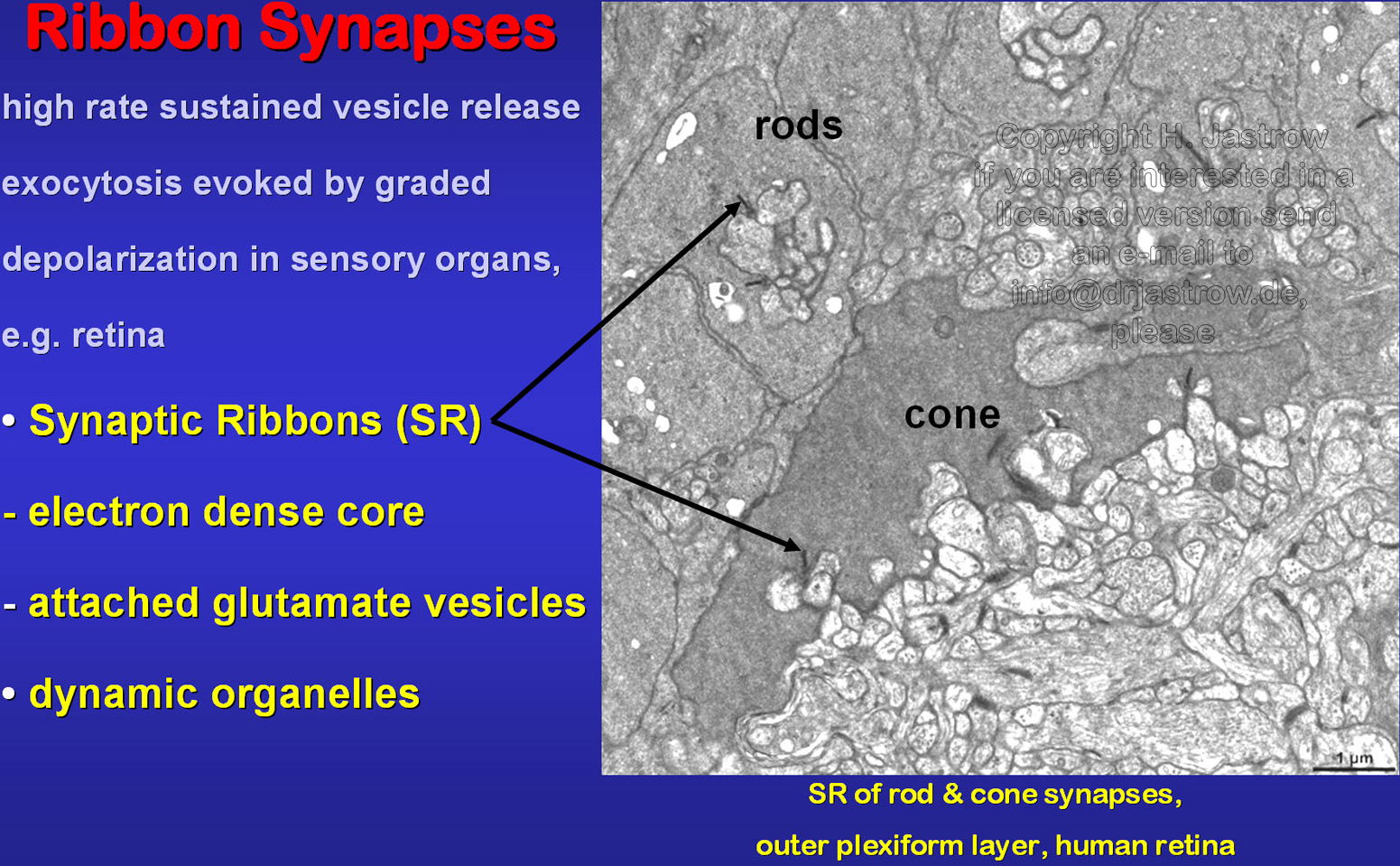 |
Fig.3
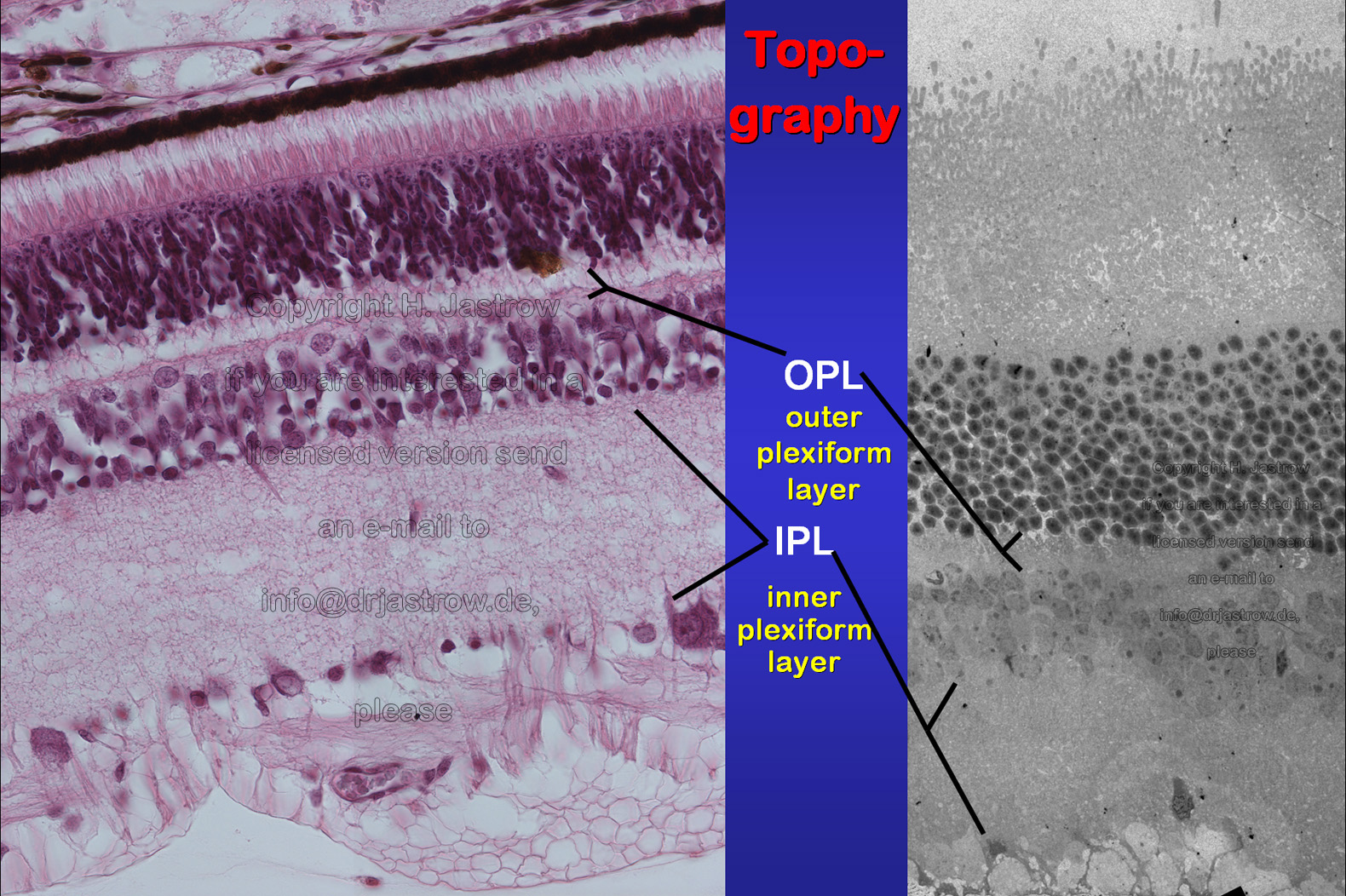 |
SR are present in rod and cone terminals of the outer
plexiform layer and in bipolar cell processes of the inner plexiform layer
of the retina (Fig.3).
Serial sections of 5 different groups of animals (Fig.4) were micrographed on the electron microscope. Nine week old animals were decapitated the end of the dark phase, others at the early and late light phase, furthers after light exposition during the dark period or chemical light pretending. Different software was used to overlay, segment and visualize synaptic ribbons in rods, cones and bipolar cells. |
Fig.4
 |
Fig.5
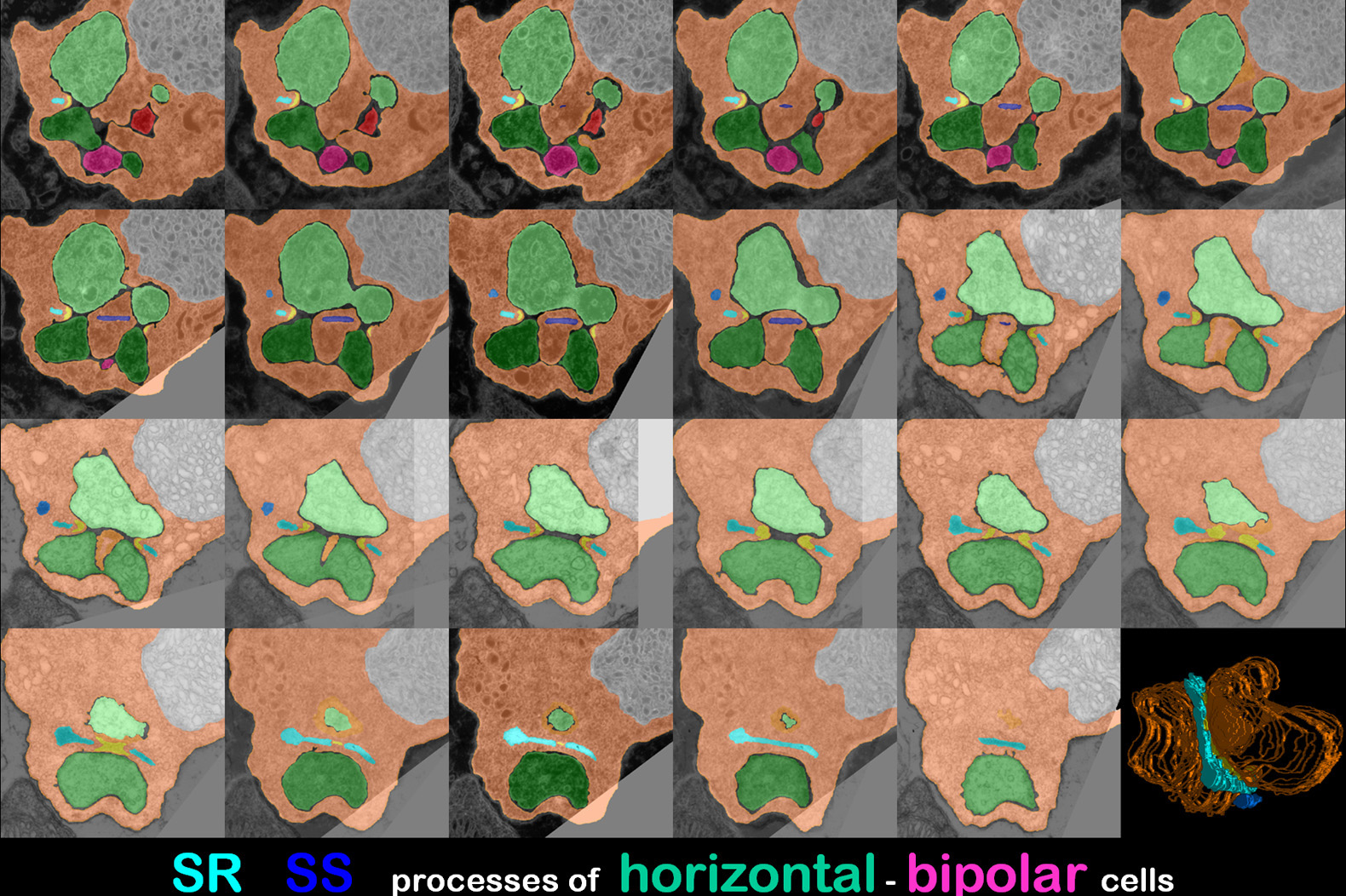 |
Thereby different colours were used for coding
ribbons and post synaptic horizontal or bipolar cell processes in serial
sections (Fig.5).
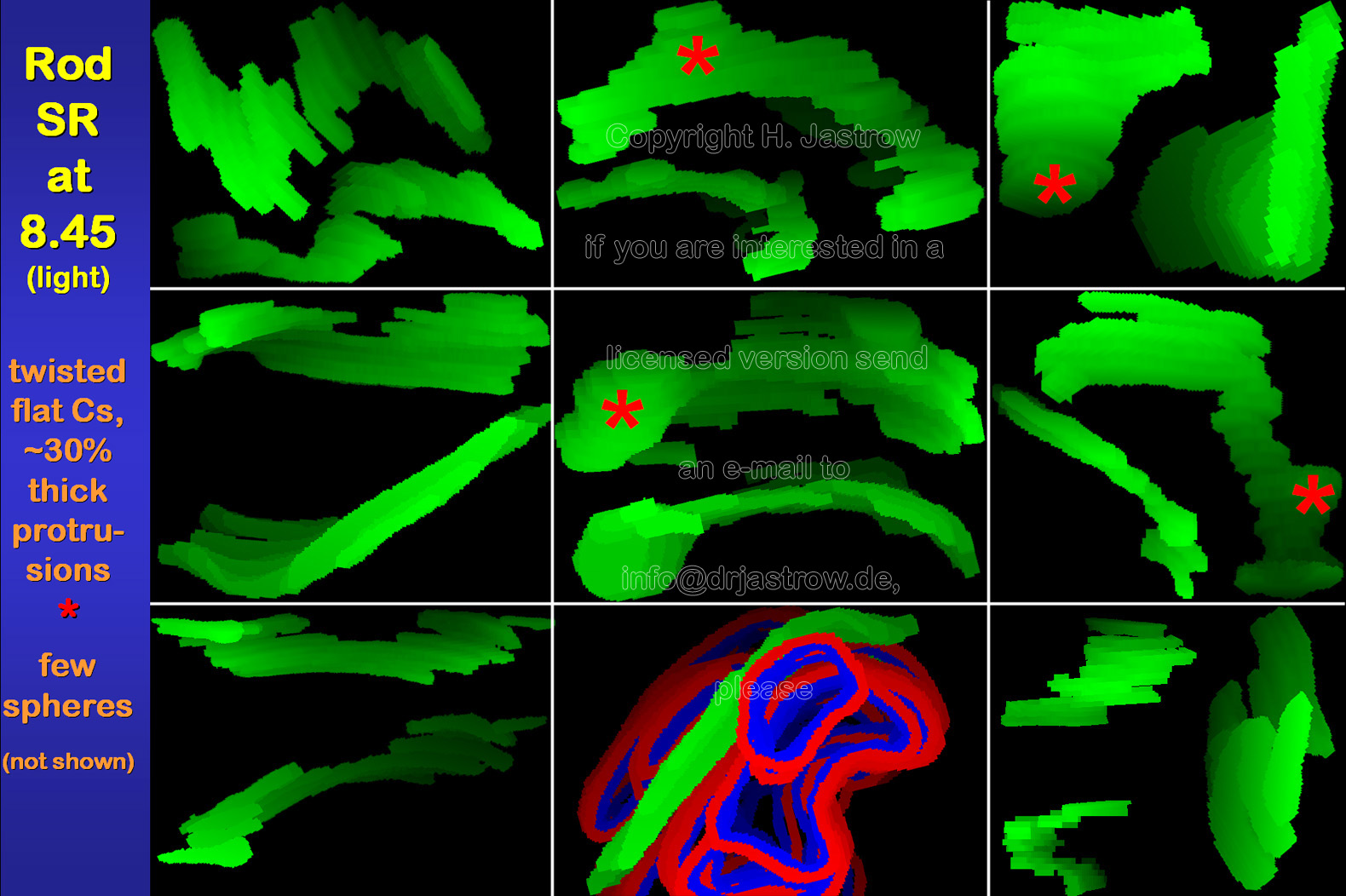
At the end of the dark phase, rod SR usually were hardly twisted boomerang like organelles with 2 major flat parallel surfaces and a thickness of about 35 nanometres (Fig.6). Mostly, they are oriented at about right angle to the presynaptic membrane in the centre between two horizontal cell terminals. |
Fig.6
 |
Fig.7
 |
About 30% of all SR show swellings mainly on
their lateral sides and not their ends at the early light phase (Fig.7).
Further some spherical organelles appear.
Besides that there is no obvious change in synaptic ribbon morphology in comparison to the previous group. Three dimensional shape of SR is not strikingly further altered at the end of the light phase (Fig.8). However, some seem to have a thinner central area. Similar to the earlier light phase, some of the organelles show protrusions and a few are spheres. |
Fig.8
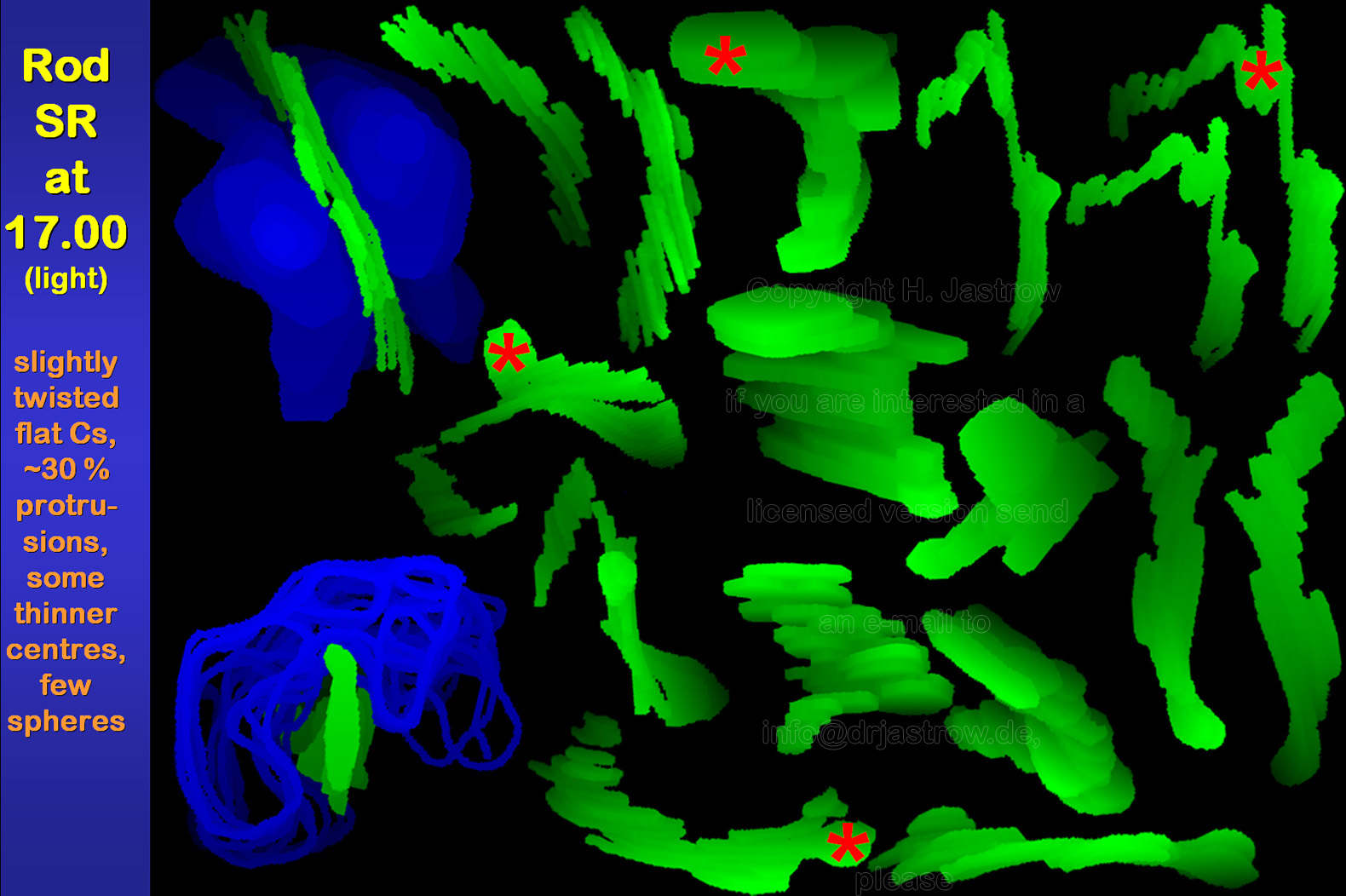 |
Fig.9 -
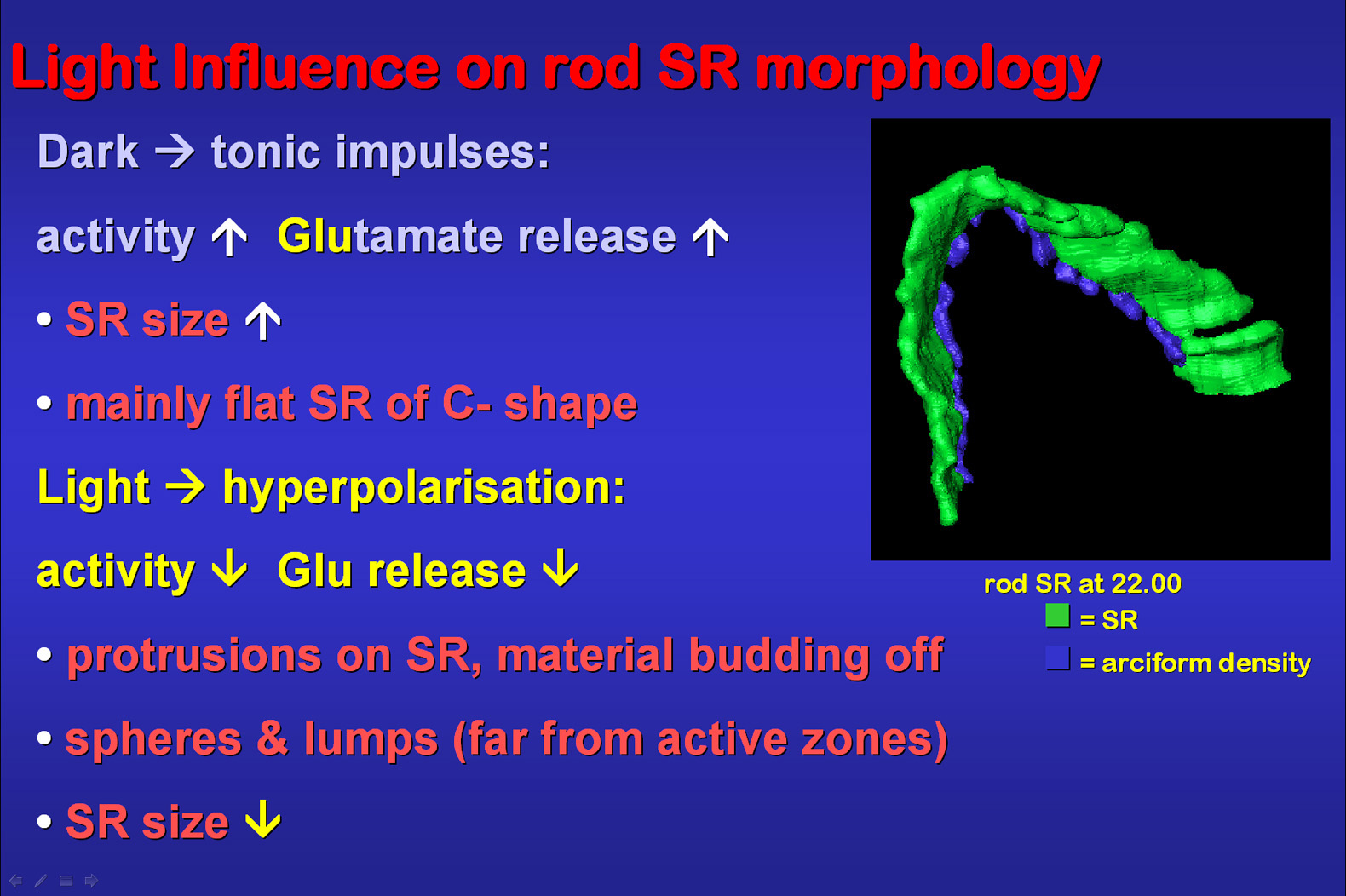 |
Fig.9: Darkness induces a maximal activity
on ribbon synapses causing a high glutamate release. In so far it makes
sense that SR are larger than under light conditions in order to offer
a maximal surface area for keeping vesicles ready for release in minimal
distance to the active zones of the synapses. Their 3D C-like shape is
optimized for the space available between the adjacent horizontal cell
processes. Light reduces glutamate release and ribbon synapse activity.
About 30% of the SR show protrusions of budding off material gathering
as spheres or irregular lumps which are seen relatively far from the active
zones. The arciform density which is immediately attached to the cell membrane
and the SR via thin proteinaceous stalks was also reconstructed in the
shown animation.
Rod bipolar synaptic ribbons are much smaller than those of the photoreceptors (Fig.10). They are flat, slightly curved plates with a sizeof about 150 x 250 nanometres (nm). Most of them are located less than 200 nm distant to the plasmalemm. However, in contrast to rod SR only some of them are directly attached to the cell membrane by thin electron dense material. |
Fig.10
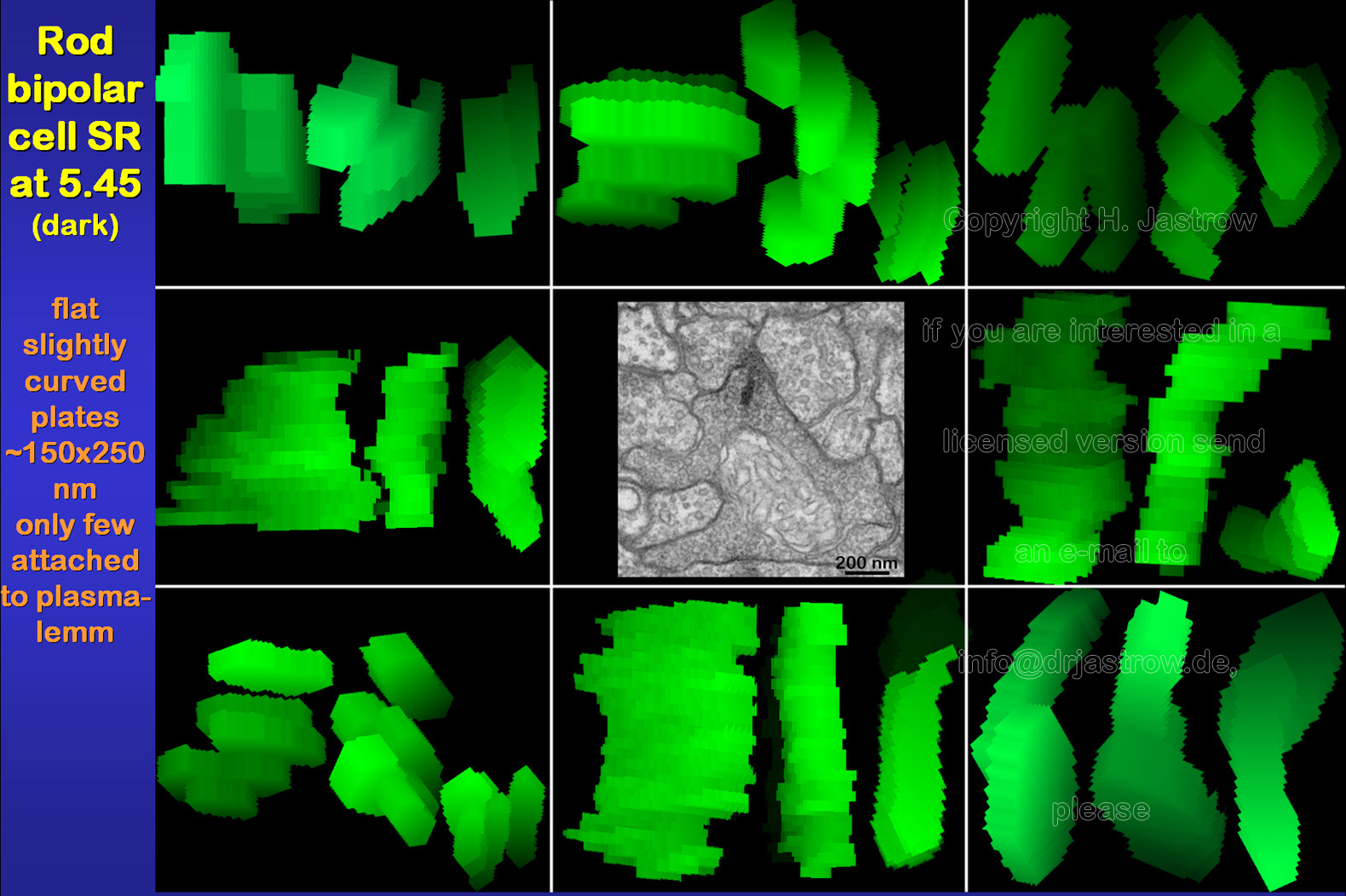 |
Fig.11
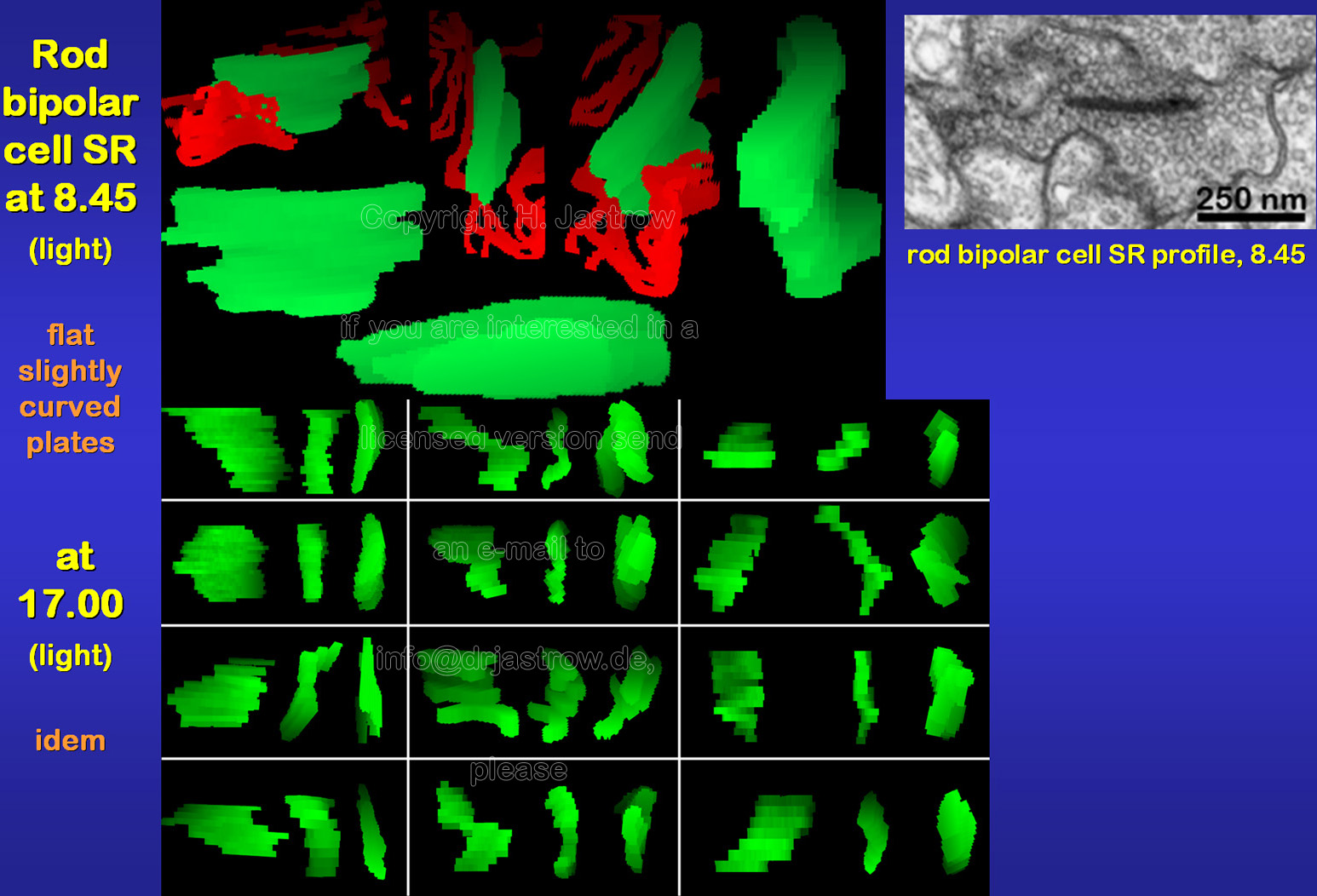 |
There is no obvious difference in morphology
of rod bipolar cell SR under different light conditions (Fig.11).
Fig.12 summarizes their general morphology: SR are small slightly curved plates with a rather constant thickness about 30 and a size about 150 x 250 nm. Most of them lie close to the cell membrane but only few are directly attached to it as also demonstrated in the animation. |
Fig.12 -
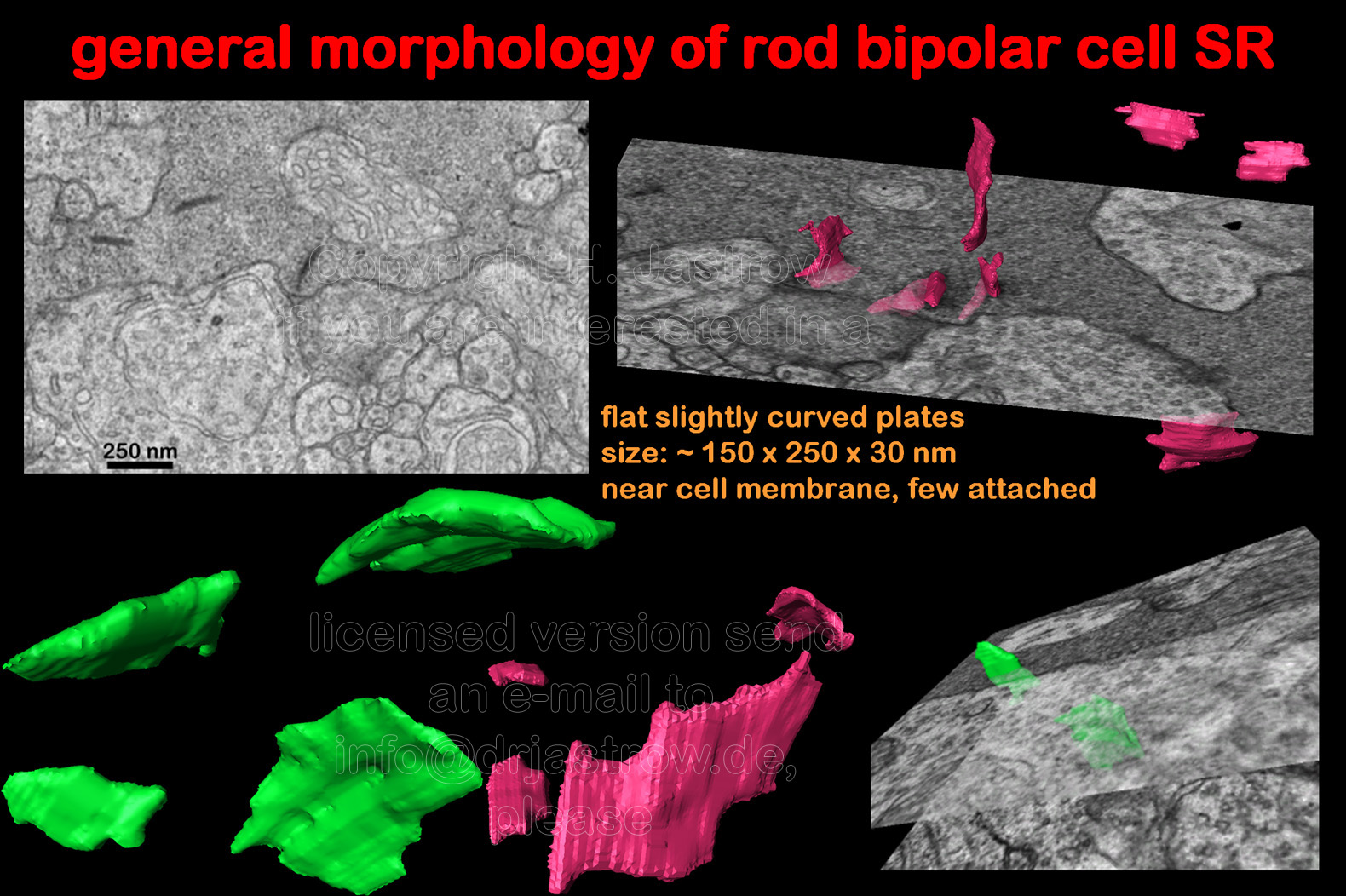 |
Fig.13 click for
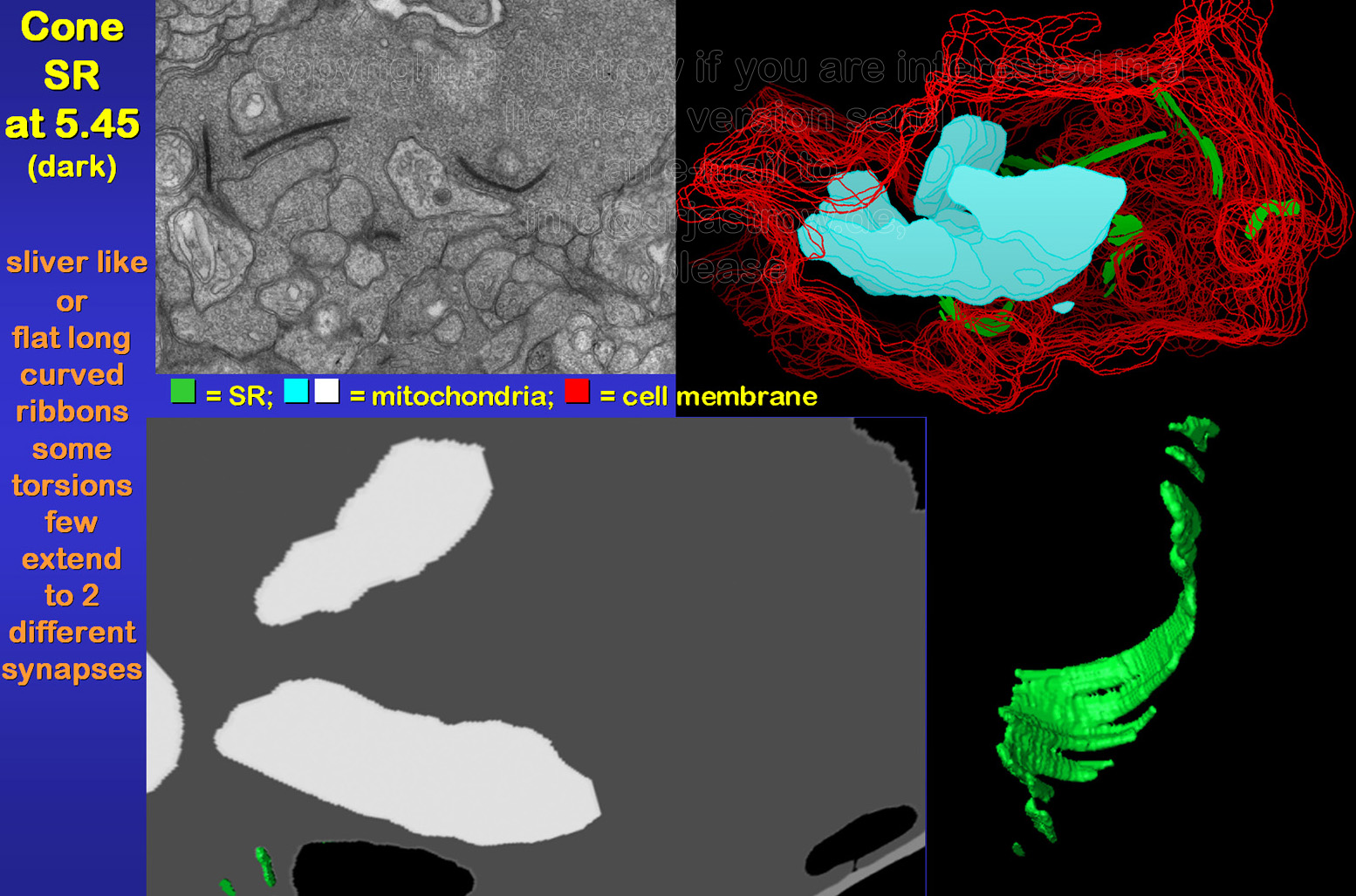 |
Cone SR are larger than those of
rods or bipolar cells in general. They may extend over 2 µm (Fig.13).
Usually 15 to over 25 of these organelles are present in a complete cone
terminal. Most of these pre-synaptic structures are flat long curved ribbons
mostly with considerable torsions as demonstrated in animation
1. They are often involved in two different synaptic complexes
as shown in animation 2. Further smaller
sliver like SR are present.
The SR reconstructed from cones at the end of the light phase were not different from those of the animals killed at 5.45 (Fig. 14). In contrast to rod SR they did not show any protrusions and spherical forms were not encountered. |
Fig.14
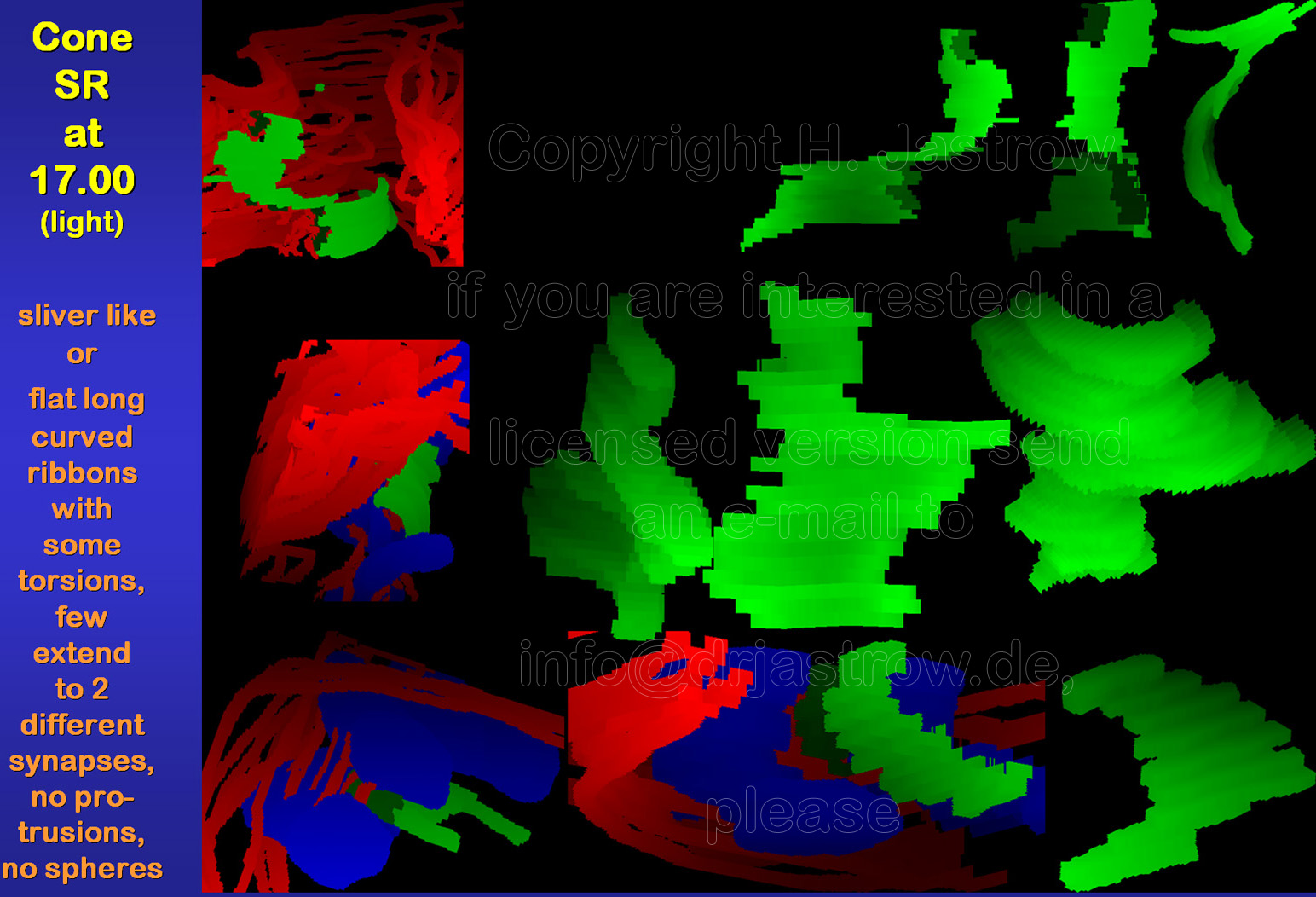 |
Fig.15 -
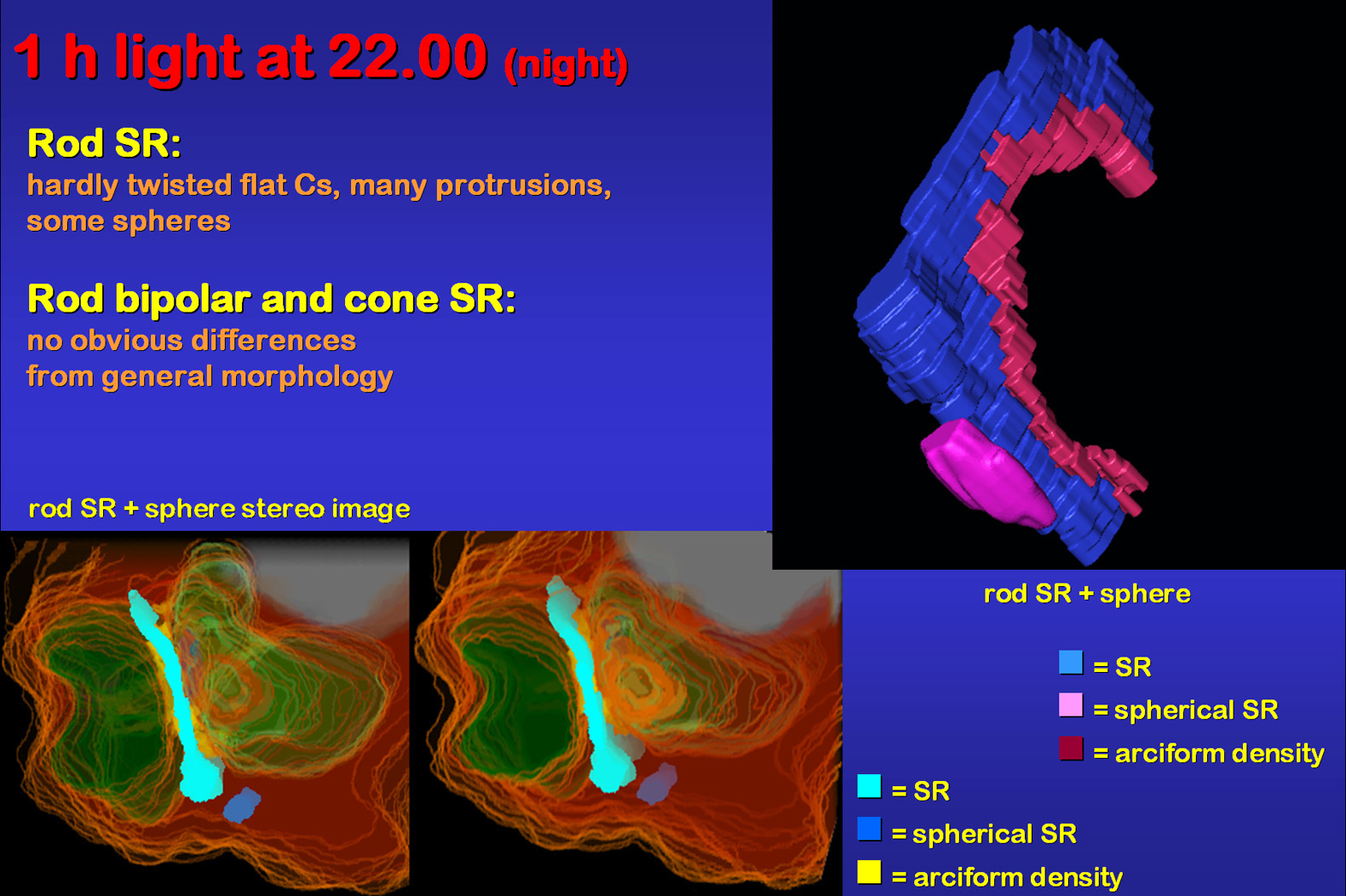 |
One hour of light during the usual dark period caused changes similar
to the light groups, i.e. protrusions of SR and spherical forms (Fig.15).
This can be seen in the animation and
the stereo image. Both of which were reconstructed from the same SR using
different software.
The removal of calcium ions from organ culture media by application of EGTA demonstrates that SR morphology in rods depends on these ions (Fig.16; Spiwoks-Becker et al. 2004). The investigated retina had never seen any real light and showed the most striking alterations of SR. As can be seen in animation1 and 2 only a very thin ribbon material bridge remains between the extreme protrusions at the endings of the ribbon. Typical C-shaped structures comprised less than 10% of all SR. From these findings we conclude that light information alters ribbon morphology via calcium ion reduction in rods. Cones were not reconstructed so far but their profiles did not show obvious alterations. |
Fig.16 click for
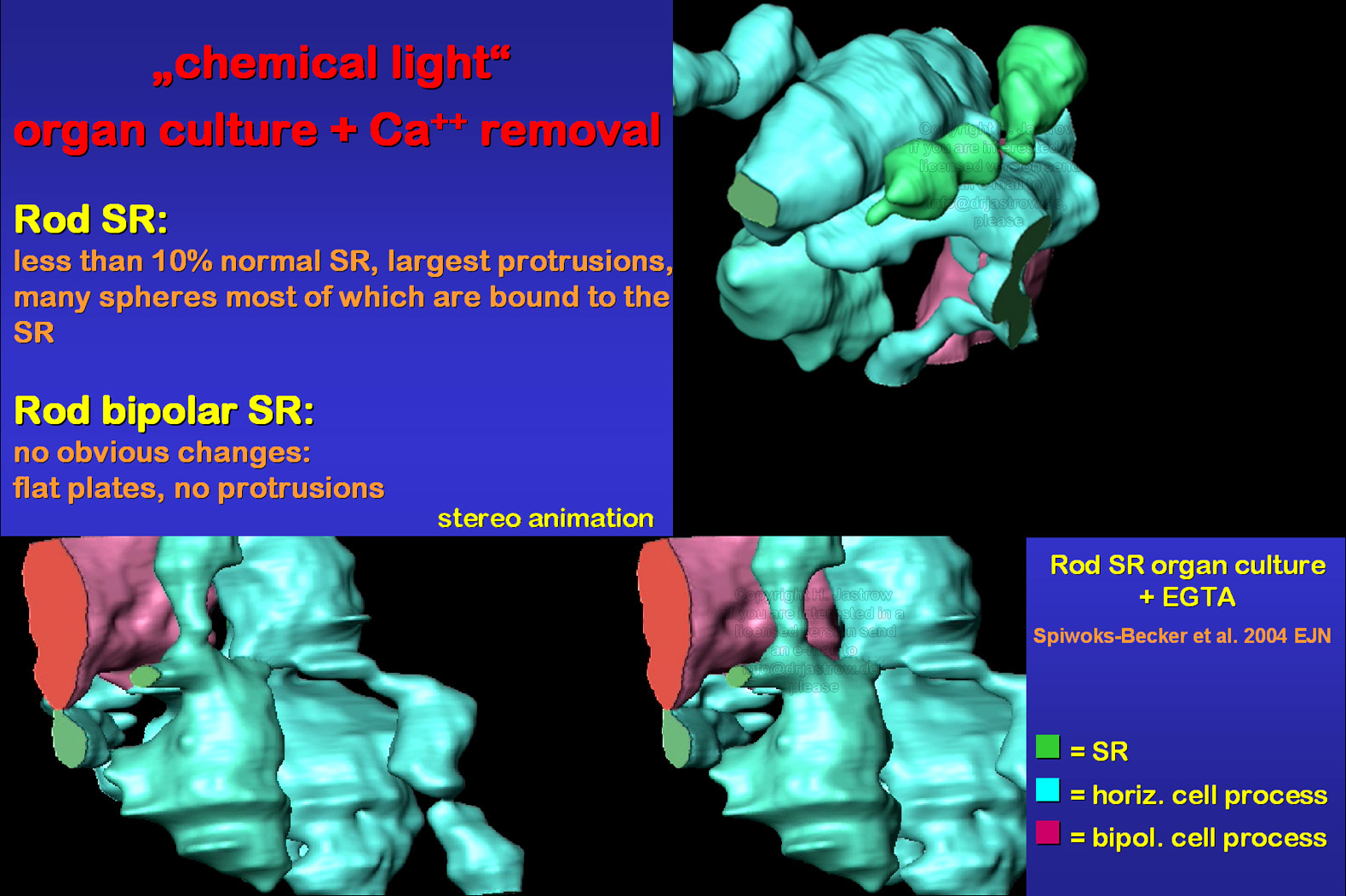 |
Fig.17
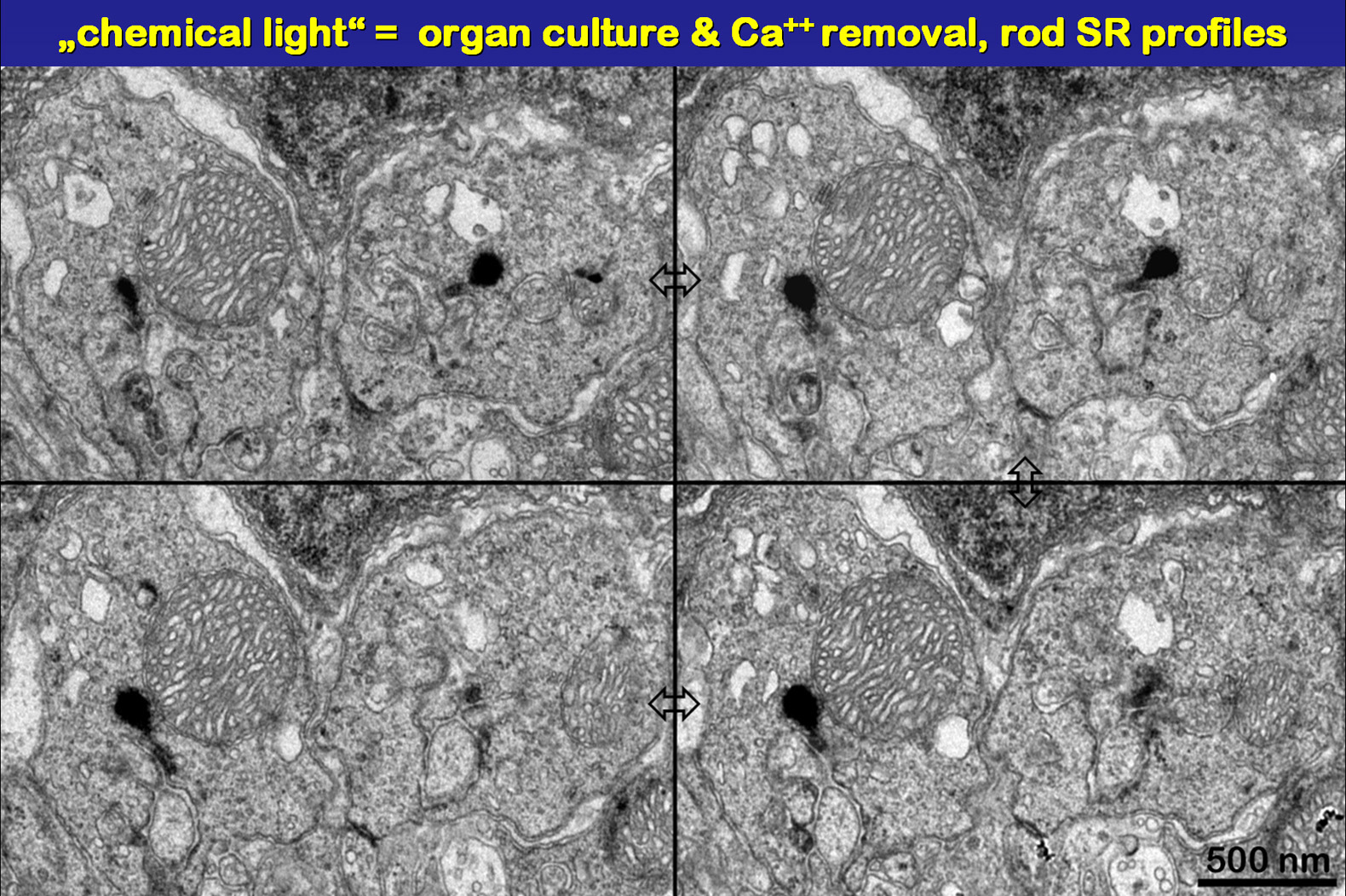 |
The serial sections in Fig.17 show profiles
of distally swollen rod SR. Note that there is hardly anything left from
the rod like basic structure attached to the arciform density.
The assumed functions of ribbons are listed in Fig.18. Most important of which are storage and refilling of vesicles as well as keeping them close to the active zones of the synapses. This is necessary for maintaining the high frequency of transmitter release typical for ribbon synapses when light is off. Note that the SR of the shown cone synapse is located exactly in the middle of the two horizontal cell terminals, that it is at right angle to the cell membrane and attached to it by fine filaments via the arciform density. The distance of the active zone of the synapse to the ready for release vesicles at the base of the SR is just a few nm. |
Fig.18
 |
Fig.19
 |
Fig.19 lists the molecular components
of SR known so far. Of special interest may be KIF3a, that as kinesion
motor, may be involved in a conveyor belt function of SR. Bassoon is localized
in the anchoring arciform density and basal SR region. Knockout of Bassoon
as performed by Dick
et al. (2003) resulted in free floating SR and severely attenuated
transmission from photoreceptor to bipolar cells.
I thank the following persons for their support and advice any you for your attention! |
Fig.20
 |